背景 #
TE介绍
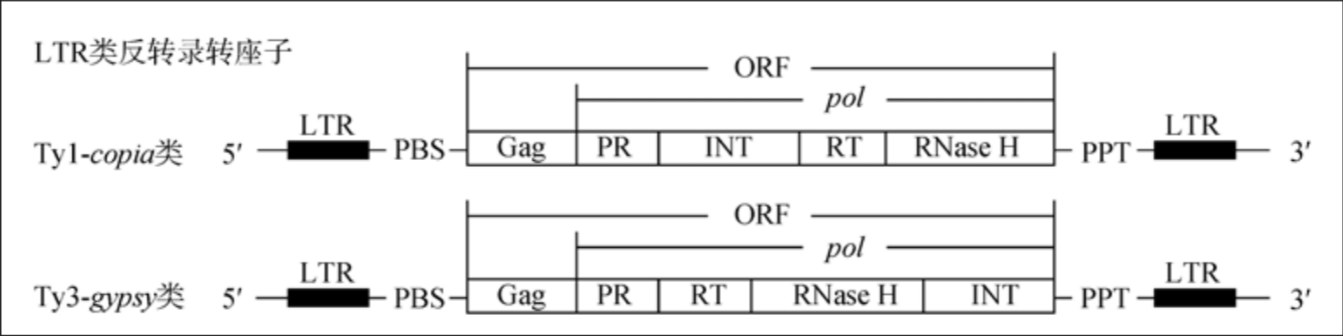
侯小改,张曦& 郭大龙.植物LTR类反转录转座子序列分析识别方法.遗传 34, 1491–1500 (2012).
- 分类及介绍: 小型遗传单位, 能够自我复制或在基因组中移动
- 逆转录转座子: 通过mRNA的逆转录功能进行转座
- DNA转座子: 代表其他所有的转座子
- 基因组多样性的主要因素, 也是解释基因组大小变化的主要因素
- 经常被表观遗传沉默, 使其无法长期大规模扩增, 之前观点认为它们可能逃脱沉默, 大规模扩增并迅速沉默
- TE对基因表达也有重要的影响, TE也可能是等位基因的主要来源
结果与讨论 #
TE content and distribution along the 21 bread wheat chromosomes #
三个小麦亚基因组的 TE 组成和染色体分布示例,在超家族层面上, 含量类似, D 亚基因组较小的原因主要是Gypsy超家族含量较少

|

|

论文里面作者还讨论了很多其他家族的情况, 并且发现了小麦特有的家族, 因为分析过程和以上区别不大这里就不展示了.
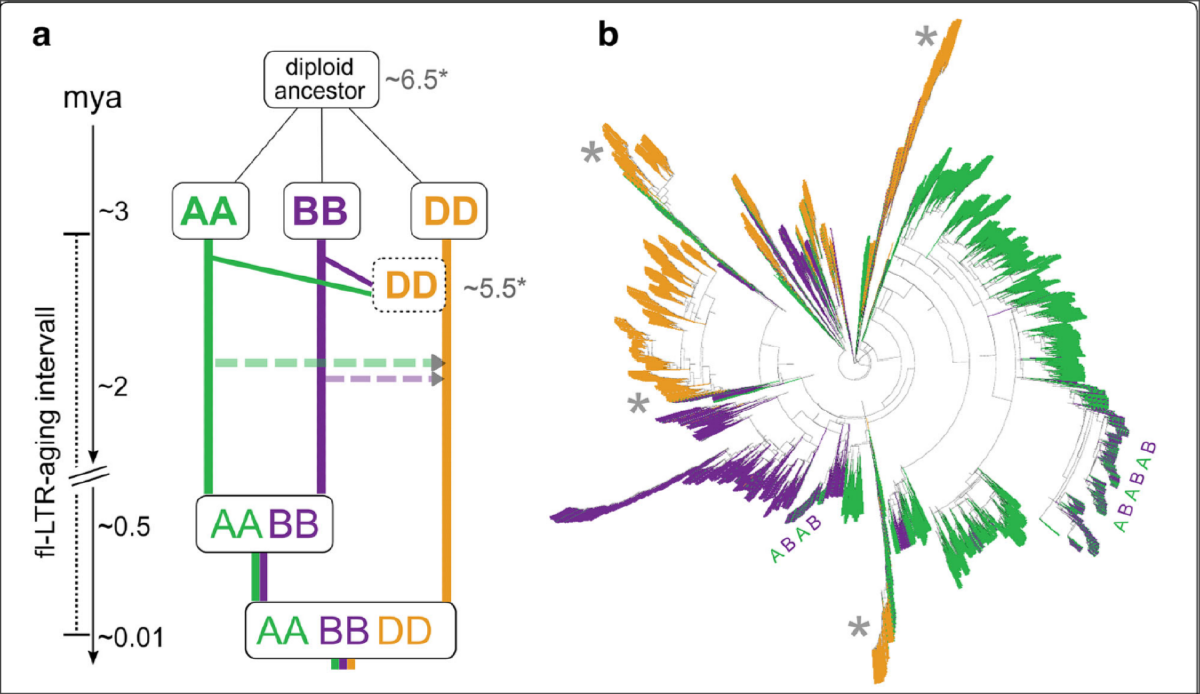
Dynamics of LTR retrotransposons from the diploid ancestors to the hexaploid #
旧的TE可能会经历多次的嵌套插入,截断, 序列突变(truncations, nested insertions, and mutations), 最终导致结构缺失难以识别, 所以完整的全长LTR(flLTR-RT)代表最近插入的TE, 作者从中国春基因组中共发现112,744个flLTR-RTs. 数量远远大于之前发表的小麦基因组, 原因可能是组装完整度的提高.
SanMiguel, P., Gaut, B.S., Tikhonov, A., Nakajima, Y.& Bennetzen, J.L.The paleontology of intergene retrotransposons of maize.Nat Genet 20, 43–45 (1998).
LTR-RTs两端的重复序列会随着时间发生一些突变, 通过计算突变率可以大致估算其插入时间, 作者计算了这些flLTR-RT的插入时间并分析了flLTR-RTs和最近插入(两端完全一样)的TE在染色体上的分布, 发现最近插入的TE并没有显著的位置偏好, 证实之前的假设, 即TE在整个染色体上以相同速率进行随机插入, 但是在末端受到选择, 以更快的速度被删除.
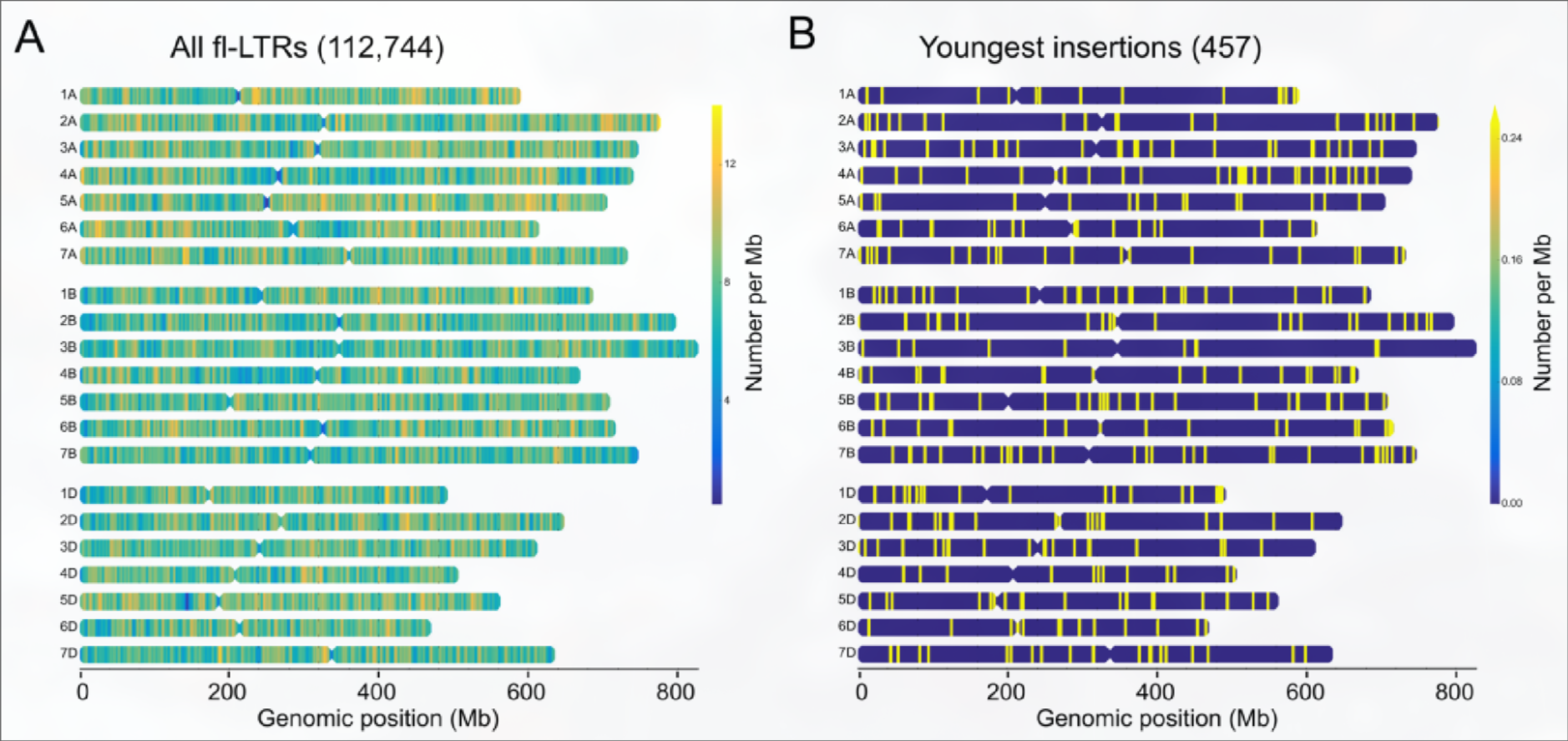
与AB相比, D亚基因组的含有更年轻的flLTR-RT.

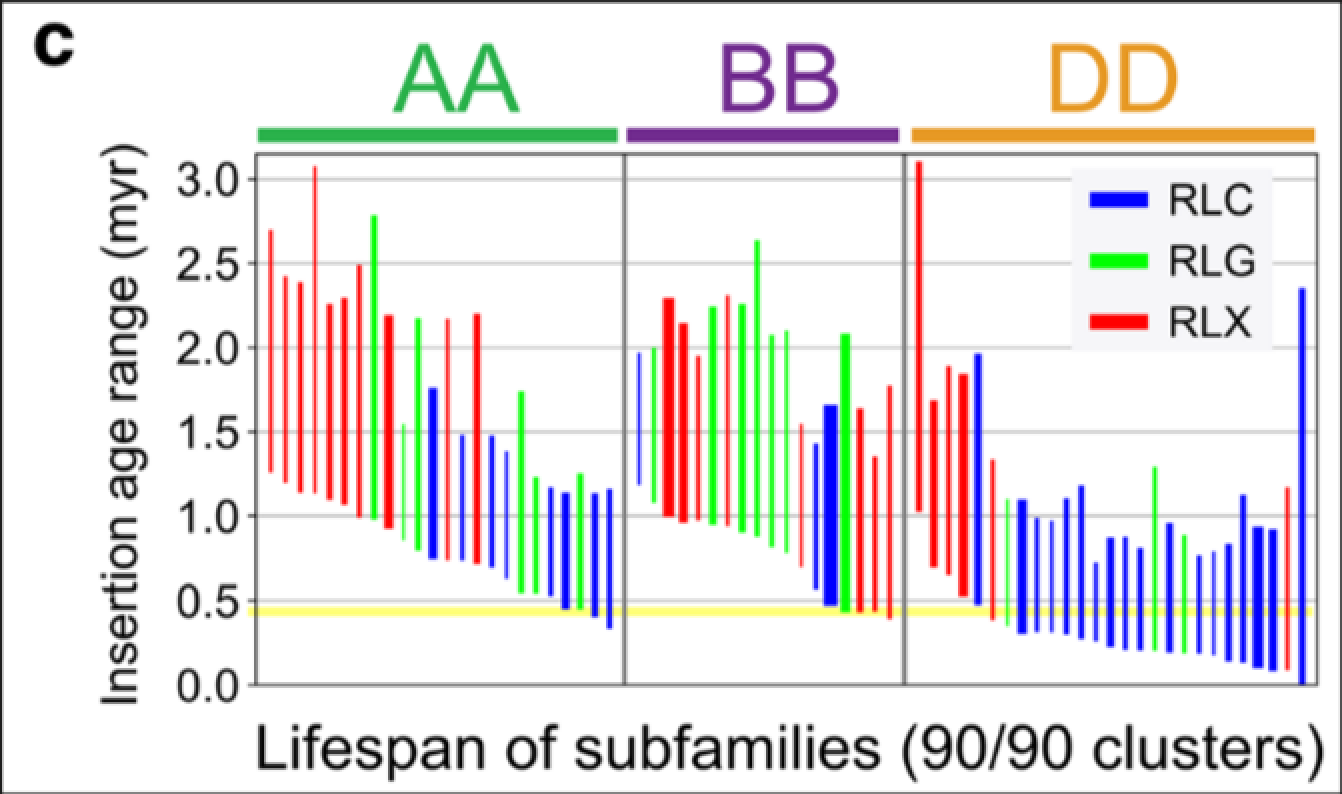
接下来是我个人认为最精彩的一部分工作, 作者对flLTR-RTs进行了不同程度的聚类分析(80/80代表80相似度,80覆盖度), 并分析聚类的簇的亚基因组偏好性. 因为90/90的参数下聚类得到结果较好, 所以后续分析是以90/90为标准进行的. 对亚基因组特异性flLTR-RTs的插入时间进行统计, ABD特异性簇的进一步分析, 粗细代表簇中含有的数量. A或B特有的簇仅扩增至0.5百万年, 与四倍体事件时间一致, D亚基因组特有的簇是最近插入的, 与六倍化事件时间一致.

|
|
作者也对最大的三个聚类簇构建了系统发育树, 大部分的扩增发生在二倍体祖先中(这些单一颜色的分支), 每棵树包含一个或多个较年轻的区域, 其中交织这分布在A和B基因组中的TE, 说明这些扩增是在四倍体中进行的, D基因组插入时间较短, 还未留下此类痕迹.

其他结果 #
- 作者统计了同源基因在三个亚基因组中的顺序和相邻基因的距离, 尽管存在大量的TE插入, 但是基因的顺序和相邻基因的距离都非常保守, 说明存在某种选择
- 分析转录起始位点上游 10 kb 和转录终止位点下游 10 kb的TE分布情况. 在超家族水平上,A、B 和 D 亚基因组在基因周围区域表现出相同的偏向组成
- 作者还利用公共转录组数据研究了基因组表达与启动子中特异TE家族的关系, 发现没有显著的关联.
|

|